
INTRODUCTION
The material presented in this chapter represents the first part of the dissertation. Three questions are asked; 1) Does evidence from the literature tend to support Bates? 2) Can an etiologieal model be developed on the basis of Bates’ and
psychophyaiological concepts which is compatible with the literature on nearsightedness? 3) Should future research be conducted on the basis of Bates* conceptions? In other words, is it time to take another look at Bates?
Previous studies of refractive change in populations of subjects will be reported first. Literature concerning five approaches to studying the etiology of myopia as well as a critique of each approach is then presented. Next Bates* ideas are considered. Recent neurophysiological evidence on vision, attention, and brain function is then reviewed in light of Bates1 model. This evidence emerges largely from the conceptions of Karl Pribram. A paychophysiologic&l model of myopia is then developed on the basis of the information presented in this chapter. The model is compared with the literature on the
etiology of myopia. The research questions are discussed and
conclusions are drawn, and the discussion and implications for future research will be presented in the final chapter of the dissertation.
STUDIES OF REFRACTIVE CHANGE
Studies of refractive error have been reported in the
literature in the past which examine variables or age, gender, dioptric level, and refractive changes over time. These methods of analysis were used in the pr&sent study. The results of the previous
research will be an additional check on the reliability of this study.
The following studies are a review of data by which researchers examined samples of individuals over the past decades. In order to determine the most pertinent questions which relate myopic behavior to test probes, the past studies will be examined. These data will provide a standard or control to judge the reliability of the present study.
In the studies of nearly 20,000 patients by Tassman (1931), Jackson (1935), and Walton (1950) the percentage of myopes,
emmetropes, and hyperopes at various ages shows that the relative frequency of hyperopes decreases over time; the emmetropes
increase over time; and the myopes first increase, then decrease, then later increase in frequency with age. Information about
individual variations in refractive error or refractive change is not indicated by this method of
analysis, but it can be concluded from examining Figure 1 that myopia does spontaneously decrease in the young adult age group.
Analysis of the trajectory of refractive error (Bucklers, 1953) in Figure 2 shows that when one individual is measured over a number of years, the slope of the trajectory for some
patlenta changes with age and that individuals seem to have their own unique rate of change. It will also be noted that some myopic patients seem to improve at various ages after the age of about twenty. Trajectory analysis provides one means for comparing refractive changes at specific age intervals.
Hofstetter (1954) in analysing the refractive status of 579 patients taken from the files of a practitioner, computed the spherical equivalent of both eyas determined at each of two
examinations. He further computed the average monthly change in this mean spherical equivalent for the twenty-one to thirty-four age group. The scatterplot for the distribution of changes is
plotted in Figure 2. This method of data analysis spotlights the differences in speed for various refractive error levels.
Examination of the plot shows little variation due to dioptric level; however, it will be noted that a number of myopic patients showed marked improvement in refractive error, i.e., they had velocity rates in the positive direction.
The next data analysis plots the changes in refractive velocity itself as a function of time. Figure 4 shows such a treatment by Slatper (1950). This method of analysis indicates that there are trends and predictive future changes; age is an important variable in the behavior of myopia; and that there is an indication of improvement in myopia in the late twenty year old population.
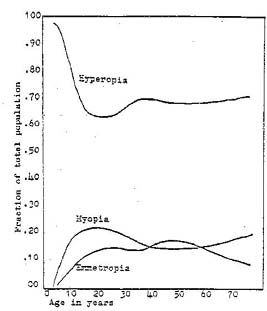
Figure 1 – Fraction of total population with indicated ametropia at various ages.
The fraction of emmetropes, hyperopes, and myopes for a total (non-clinical) population is plotted with respect to age. Myopia increased in this population until about age eighteen and then diminished steadily until about age forty-five when the fraction of human adults having myopia again increased. Adapted from Tassman, 1932.

Figure 2. Trajectory of the refractive error at indicated age.
Data was plotted from the records of patients who were examined over a period of years. Each line represents one patient. The dioptric value of the refractive error is plotted with respect to the age at the time of examination. Note the apparent decrease in myopia for some patients. Adapted from Bucklers, 1953.
Four different methods of analysis have been presented showing the sensitivity of the probe for refractive information. The studies show that age is an important variable in refractive
change hut that level of refractive error is not so apparently myopic related. In all the studies there were
indications that patients in their twenties and thirties often
improve spontaneously.
The present sample of myopic patients will be analyzed by these methods and the results will be compared with the results of other studies.
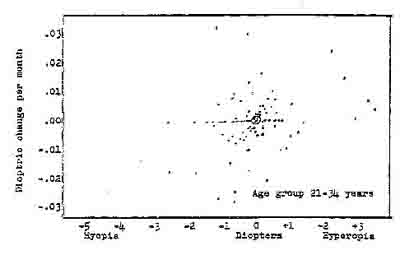
Figure 3. Dioptric activity as a function of refractive error.
Within this population from Hoffstetter, 1954) there appears to be a little correlation, that is interrelatedness, between refractive errors and monthly rate of change of refractive errors. But note that some myopes (upper left-hand quadrant) appears to decrease. Each dot represents the average of two eyes. The average velocity for the present study is plotted as indicated by o.
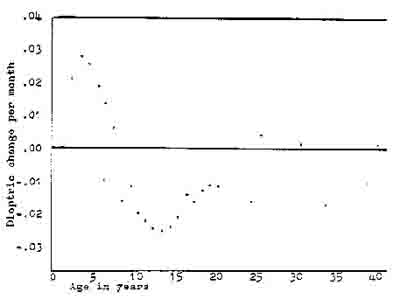
Figure 4. Dioptric velocity according to age.
Each point represents the dioptric change per month averaged for a number of patients at a given age. The sign, plus or minus, of the dioptric velocity indicates the direction of change towards or”away from myopia, but the magnitude is the magnitude of the change par month. The age group before age seven, appears to be increasing in hypercpia but at a declining rate. After age seven myopia increases until about age
twenty-five when it begins to decrease. The rate of change in the myopic direction increases after the age of seven, peaks at age fourteen, and diminishes until the mid-twenties when it reverses again away frcm myopia. The average rate of change for the average age in the present study is also plotted indicated as o. Adapted from Slataper, 1950.
ETIOLOGY OF MYOPIA
There bas been a wide range of efforts to determine various behavioral and physical characteristics of people with refractive problems. These efforts to determine the etiology of myopia have resulted in five general approaches to the subject (Kelley, 1958):
- The Genetic Theory – Myopia is an inherited condition.
- The Nutritional Metabolic Theory – Myopia is a product of the diet and/or metabolic factors.
- The Conditions of Use Theory – Myopia is the result of the excessive use of eyes for near-point tasks.
- The Normal Biological Variation Theory – Myopia and hyperopia are, in most cases, only the reflection or normal variation in the length of the eyeball and the strength of the eye’s optical elements.
- The Psychological, Personality, Emotion Theory – Myopia and hyperopia are symptoms of emotional habit attitudes, due to repressed emotion, as a direct defense against anxiety or as symbolic wish fulfillment,
self-punishment, etc.
Genetic Theory: There are a large number of variables which make up the optical anatomy or the refractive accuracy of eyes (Figure 5). These include the curvature of the optical elements (surfaces of the cornea and the lens); the physical length (axial length) of the eyeball from the cornea to the retina; the positioning of the optical elements; the thickness of the optical elements; and the index of refraction of the cornea, lens, end media or the eye. Each of these variables might be influenced genetically. Until we know the nature of this influence on each of these
components of the eye, we cannot know the genetic implications in the incidence of refractive error.
A representative statement of this position has been made by Baldwin (1964):
It is known that myopia can be congenital without direct hereditary influence. It is also known that the incidence of both hyperopia and of myopia are influenced but not determined solely by heredity. These facts are demonstrable by studies of the
refractive status of twins; studies of family pedigrees; and by noting the incidence of hyperopia and of myopia associated with genetic syndrones.
For a detailed review of the many investigations in this area the textbooks of Frankeschetti (1953), Francois (1961), and Wardenburg (1962).
Some people do seem especially susceptible to conditions which cause refractive error, and the possiblity exists that these differences in people may be due to inherited
constitutional factors (Henderson, 1934). The studies of identical twins by Hofstetter and Rife (1953), and Meyer~Schwickerath (1949) show a strong correlation among monosygotic twins but not to the degree required for a conclusive genetic theory.
Young (1956) concluded that it is unlikely that refractive error is determined primarily by hereditary factors.
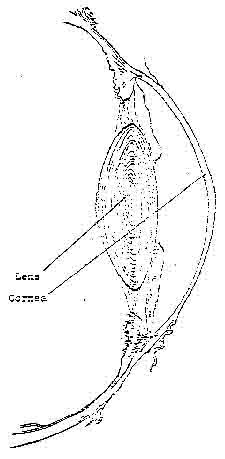
Figure 5. Anterior section of the human eye (schematic).
Drawing shows the optical elements which contribute to the refractive condition of the eye.
Young (1956) concluded that it is unlikely that refractive error is determined primarily by hereditary factors.
Nutritional and Metabolic Theory: Price {1939) in a cross-cultural study, observed that myopia was less prevalent in those cultures which maintained their traditional diets. He attributed
degenerative effects to the paucity of vitamins in cultures relying heavily on commercially prepared fooda. He correlated the
incidenee of myopia with dental caries, as did HIrsch and Levin (1973). Young (1969) found a 59 percent incidence of myopia among sixth grade Eskimo school children, but only a 5 percent incidence in their parents, and none at all among their grandparents. This study yields evidence which counters the genetic theory of myopia, but it does not indicate that nutritional factors are necessarily the determining factor. Other cultural changes, such as the transition to electrical lighting and mass western educational methods also must be considered.
Livingston (1964), Reed (1947), Balcet (1948), and Smith (1951) found an increase in the incidence and degree of myopia among war prisoners and attributed this to malnutrition. McLaren (1960) observed a high incidence of myopia among children of the Gogo tribe in Tanganika who had suffered from two years of severe famine. Despite this finding he concluded from this and other
studies in his publication, Malnutrition and the Eye, that:
“an early and almost constant feature of nutritional deficiency is retardation of body growth. The eye is quite resistant to this influence, however.”
The war prisoners and the Gogo and the Eskimo children were also Subjected to other stresses of a serious and unusual nature; thus, malnutrition was not the only possible related variable.
Wood (1927), Feldman (1950), and Law (1934) found that young myopes tended to have depressed blood calcium compared with
controls. Knapp (1939), Feldman (1950), and Stansbury (1948) did not find, however, that administration of calcium and vitamins (especially vitamin D) arrested the progress of myopia. There may be other reasons than diet for the children’s low blood calcium.
Some endocrinological and metabolic relationships have been investigated. Bothem {1931} determined that the basal metabolism rate in myopes was somewhat below normal, indicating hypofunction of the thyroid, Linder (1939) concluded that the greater
metabolic demand of near work (reading) produced abnormal composition of nourishing fluidsf softening the sclera. DeVreis (1950) also concluded that low thyroid was implicated in myopia. Lijo (1939) implicated the para intermedia of the pituitary because of the association of pigment epithelium changes of the retina. Filatov (1940) also postulated hormonal influence in myopia and reported increased acuity in subjects given 10 percent saline
intravenously. Borrello (1951) implicated hyposecretion of the adrenal cortex. Tasaka (1951 reported labile galvanic skin response in myopes. Temporary increases in myopia from one to six diopters due to the side effects of certain drugs have been found by several investigators (Dellaporta, 1942; Kramb, 1951; Stern, 1955; Back, 1956; Halpern, 1959; Galin, 1962). The drugs implicated
included sulfanomides, corticotropin, carbonic anhydrase inhibitors, and autonomic blocking agents. The condition disappears within a few days after cessation of use of the drug.
Conditions of Use Theory: Studies have been made which attempt
to show that there is a positive correlation between myopia and neap work, reading, profession, urban dwelling (vs. rural), and intelligence (Borish, 1954; Kelly, 1956, 1971; Baldwin, 1961; Young, 1966). These studies generally, but not exclusively, showed a positive correlation, but it has remained difficult to say whether the conditions of civilization caused the myopia or whether myopes gravitate toward civilized conditions of living.
The facts supporting the use theory relate to the increase of myopia from a very low one percent in the under five-year-old group, 8.6 percent in the five to ten year-old group, 19 percent in the 10-15 year group, and 24.17 percent in the 15-20 year-old group (Tassman, 1932). That there are other factors in the school environment which might be more relevant to the incidence of myopia are evident from a work in progress by Streff (1974). He indicates a very significant decrease in myopia in an
elementary school which is designed to reduce stress and anxiety as compared with a control school.
The incidence of myopia is markedly lower among peasants or farmers th*n among highly educated and professional people (Kephart, 1953). But again, other factors than near work may be causal agents, including the combined predilection towards reading and less active pursuits with an undetermined third
factor which is also related to myopia {Baldwin, 1964).
Young (1961) confined monkeys in chairs enclosed in plywood boxes, limiting their far point vision to 18 inches. The animals were kept confined nearly full time for the period of one year,
Two control groups were also measured. One group was not
confined; the other was confined to chairs in their cages. Young found an increase in myopia of 1.10 diopter for the experimental monkeys as compared with an increase of 0.19 diopter for the animals kept in chairs but not in boxes.
Young also studied three levels of illumination with caged and hooded monkeys, and found that the very low level lighting produced an increase in myopia of 0.124 diopter, the animals in the medium level showed a 0.75 diopter change but the high level of illumination was associated with a 0.25 diopter increase. He concluded as did Lucliah (1939), in an early study on humans, that light levels were an important factor in the development of myopia. Young (1966) also found that wild monkeys had a
significantly lower incidence than the laboratory monkeys. Again, because of the many uncontrolled factors which exist in such studies, the specific causal elements are not clear.
Despite the existence of these many studies, very little in the field is actually established. The research is marked by
inconsistencies and much confusion. Because of the lack of
conclusion and the feeling that little could be done about it, the causes of refractive error are scarcely mentioned in educational experiences and the students are left with the impression that, like Topsy, refractive conditions “just growed.”
Normal Biological Variation Theory: This view states that
refractive errors are due to normal variations present in all
biological systems such as height, weight, finger length, leg length, etc. It height, for example, is measured over the total sample of adults in the world, and if this data is displayed in a graph, the curve would have a bell-like shape. This is referred to as a normal curve. Normal curves are descriptive of situations which have random causal factors.
However, if refractive errors are plotted against the number of people having a particular prescription, the resulting curve is not bell shaped. The peak, which is slightly farsighted, shows a heavy excess. Far more people are essentially
emmetropic than would be expected if refraction were due to random growth factors. Some 75 percent of the population have
essentially no refractive error (Sorsby, 1964). If the components of refraction are plotted individually against the number of eyes in the population, each component does seem to have a norm distribution. This has led to the conclusion that refractive emmetropia is produced by some active mechanism which correlates the various ocular components leading to clear focus at distance. This mechanism is called emmetropization. In cases of
refractive error (ametropia) it is not the ocular components but the correlation of these components which is at fault. The
mechanism which produces this coordination is considered to be
automatic. Sorsby (1967) has stated:
There is no free association of the axial length and cornea. There is instead a highly coordinated
mechanism involving the correlation of four normally
varying components: axial length, corneal power, lens power, and depth of the anterior chamber. The
characteristic of the eye as an optical system is its marked correlation of these four components. It is this correlation which generally produces emmetropic and near emmetropic eyes. Generally the abnormal component is the axial length.
In 90 percent of the cases the axial length does tend to be short in farsightedness (Sorsby, 1967).
The mechanism producing self-focusing in the eye is
considered to be the possible result or regulation through feedback between the fovea and cortical and subcortical control systems of the brain (Van Alphen, 1967).
Psychological, Personality, Emotion Theory: Kelly (1971), in a clinical but not experimental observation, described refractive error in Reichian body energy concepts. Using Bates’ idea that myopia was caused by the co-contraction of the oblique muscles or the eyes, he assumed that this related to a flexor action and that, therefore, the energy flow is inward, contractive, toward the self. Hyperopia is caused by the rectus muscles and related to a kind of extension attitude, the reverse of myopia, i.e., outward, expansive, away from the self. The armoring posture of the myope is an attitude of blocking the feeling and expression of fear, and that of the hyperope is blocked rage.
Again, from clinical observation, Kelly described basic behaviorial and personality characteristics of myopes. He directed his attention to body structure and function and
described the appearance of the eyes. The myope tended to be:
Inward, introjective, self-oriented, shy and
withdrawn as a child t “good1” in school, child-hood temper tantrums rare, stubborn, emotionally
inflexible, more at home with self, uncomfortable
with others, often “off in day dreams”, subvocal thought, comfortable with eyes closed, and
retreats from visual perception inward. (p. 48)
In the myope, structure and function of the body tended to be:
Underactive, often sedentary, body soft,
sometimes flabby, throat, high chest, jaws, back of neck, scalp – chronically tense, jaw rotated
forward, forehead back, shoulders forward,
disconnected from feet up in the head, centered but poorly grounded, voice often breathy or husky, Becomes hoarse easily, chest often depressed, breathing blocks on inspiration, fearful, blocking terror. (p. 49)
Kelly described myopic eyes as:
usually large-looking, open, very little local armoring around eyes, squint, fusion problems not common, unusual to have pain, eye headache, eyes lack sparkle, brightness, eyes lock down or away in anger. {p. 49)
Kelly was quick to state that these are the deeper layers of the myopic traits. Such fundamental armoring patterns are often
covered over by more superficial compensating patterns of
behavior and armoring, with maturity and experience covering to some extent the deeper tendencies:
Although the myopic posture is fearful, it is not often yielding or acquiescent. There is stubbornness and determination expressed in the stiff neck and tense jaw. It expresses a layer of anger present in most myopes that must be freed before the more fundamental fearfulness becomes accessible. Occasionally an angry myope ia found, with a high stiff chest. This ahger is superficial, easily accessible compared with the deep fear underneath. (p. 53)
Lanyon and Giddings (1974) grouped the clinical observation and speculation of others concerning myopic psychological
characteristics. They supplied a list which includes such descriptive phrases as:
shy, introverted, socially awkward, having relatively few friends, tending toward
dogmatism and self-centered personality
development. (p. 273-274)
The use of psychological testing of myopes in comparison with normals and with hyperopes has generally supported the observations of Kelley, et. al. Using the Bernreuter Personality Inventory, Hull (1948) found myopic college students to be more introverted. Inertia, over-control of the emotions,
disinclination for motor movement, inhibited disposition, and strong need for social dominance were indicated for myopes in a study by Schapero and Hirsch (1952) using the Guilford-Martin
Temperament Test. Rosanes (1966) interpreted results from the Rorschach test to demonstrate that myopes tend to have a high tolerance for anxiety; exhibit a “wait and sae” attitude in stress situations; be cautious, doubting and compliant; and avoid confrontation. Young (1955) supported the hypothesis that distinct personality differences do exist between myopes and non~myopes. Myopic students scored significantly lower in need for exhibition and need Tor change on the Edwards Personal Preference Schedule, Stevens and Wolff (1965) measured the
cognitive style of myopes as tending to pay attention to the
differences between things rather than looking for their similarities.
The above studies were correlational, relying on univariant techniques. However, Young, Singer, and Foster (1975) changed the analysis from “t” tests and analysis of variance to stepwise
discriminant analysis, followed by a factor analysis of the
variables selected on the discriminant analysis. They examined 163 male myopes and 160 male non-myopea entering a military
institute, using College Entrance Examination Scores; the Alport, Vernon, Lendzey Study of Values Test (SOV); the Edwards Personal Blank (SVIB). By their methods they could correctly classify 76 percent of their population. Consistent with the above studies, the myopes preferred high statust academically oriented
occupations, and those which appeal to an introverted, accepting
person.
Palmer (1966) examined the relationship between visual acuity and personality, It is well known that the error of
optical refraction is not in an absolute relationship with visual acuity, and that although average visual acuity does increase in a regular way with refractive error in myopia, there is a spread of acuities for any refractive error. For example, a one diopter myope usually has 20/50 acuity, but people with one diopter myopia could range between 20/50 to 20/200 (Kirsch, 1945). This is
generally known in the vision-care professions, but little notice is paid to these individual variations. Palmer found that poor acuity subjects (20/200 or worse) have more inward orientation and are less sensitive to external stimuli than the high acuity (20/20 – 20/40 group, Palmer concluded that myopia develops as a result of attempts to reduce one’s contact with the external environment:
For some Individuals, unrestricted visual imput leads to painfully high levels of excitement, and…these individuals develop a myopic norm of vision or other visual impediment as a means of “gating,” controlling or reducing stimulus input, to avoid being overwhelmed with large quantities of unmastared excitation. (p. 370)
Zeiger (1976} examined the relationship between personality and vision in terms of both the refractive error and the visual acuity. She attempted to differentiate myopes as to
characteristic style, mode or being in the world. Her data on college students revealed that there were not significant differences between emmetropes and myopes when all myopes from low to high were taken together. When she divided the low, moderatet and nigh refraction myopes, and the high, medium, and low acuity subjects, she found significant differences for the students with moderate levels of acuity and refractive error. Applying a stepwise discriminant analysis to her data, she was able to predict 86.5 percent of the moderate myopes. Moderate myopes were seen to view the world as repugnant; they reject it and attempt to withdraw from it by creating a conceptual buffer arid staying internal. The world view of the low myope is like that of the moderate myopes, only low myopes show an even stronger aversion to the world. Their coping strategy is to be actively alert and active in challenging the noxious and irrational world.
High myopes looked generally more like emmetropes than did moderate or low myopes, implying a successful defense. Zeiger presents a very interesting conclusion:
The low and moderate myopes show the problems and attitudes myopes nave and the high myopes show solutions that myopea have. (p 86)
This review of the possible causal elements which might contribute to refractive error has shown a wide variety of approaches to the problem. In moat cases the results remain inconclusive. There are isolated facts scattered here and there, but there are no final conclusions. One factor does remain clear, however. The evidence existing in the scientific
literature, while not mentioning Bates except in a very few
psychological studies, generally does not contradict his thinking, and in some cases offers direct support. This will be shown in the next section, which develops a model of brain function compatible with much of what has been presented in this review.
SUMMARY OF BATES’ CONCEPTS OF ETIOLOGY OF MYOPIA
Bates made some important observations a about the
psychophysiological factors which contribute to myopia. An
inspection of his ideas reveals a holistic approach which is
surprisingly contemporary in its tone. The following summary is based on his 1920 publlication, The Cure of Imperfect Sight by
Treatment Without Glasses.[1]
Bates (1920) believed that vision problems are most likely functional responses to an unhealthy environment. These
become long-term habitual responses to stress and are ingrained in the behavior of the visual-mental system.
He was aware that soma conditions imposed on people living in modern cultures are unhealthy. In education, for example, where children are required to sit for hours at a time,
conditions are produced whloh lead to “mental strain” (p. 256). Contributing to this strain is the influence of teachers who are “too often themselves nervous and irritable” (p. 256). Bates felt that the process of learning to learn was the
purpose of education, but the emphasis in the school is too often upon grades and prizes. When the information in schools is
presented in uninteresting, non-relevant ways, when the object of auch prolonged attention has no inherent motivating interest, the student must develop a means of controlling the mind and body, compelling each by will to do its master’s bidding. Some
children developing under these conditions can endure better than others, but some cannot stand the strain; “thus, the schools become the hotbed not only of myopia, but of all other errors of refraction.” (p. 256)
Modern civilization forces individuals to exist under
continual strain, worried, lost in thought, reviewing memoirs of past experiences which filter out present experience. It is only within the last decade that popular attention has begun to focus on the production of stress-related illnesses, a
relationship which Bates pointed out over six decades ago in
conjunction with his studies of refractive anomaly.
Refractive errors are produced by a strain or effort to see. But for Bates the act of vision is passive, just as taste and touch are pasivve. The letters on a visual test card are there, perfectly distinct, just waiting to be recognized. But when we make an effort, when we try to force the eye to see, the effort sends impulses to the eye and to the extraocular muscles which tense abnormally and distort the shape of the eyeball.
At the same time the eyeball loses its ability to aim and coordinate its movement as intended. Bates felt that the visual impairment is seldom due to a faulty construction of the eyeball itself, but rather to more central errors of functioning. The basic problem lies in “wrong habits of thought” (p. 106}. Our culture, our activities, lead to the faulty mental habits which become normal ways of operating in the world. Patients must be taught to let the mind rest, to let the eyes become passive robots.
Only then will the visual system respond naturally and
spontaneously to the exigencies of the environment.
Bates pinpointed the cause of refractive errors in the way the eyes are used, and hypothesized that it is in one’s mental habits and attitudes that the fault lies. He related vision problems ^o temporary and habitual emotional attitudes. The anxieties of fear, of lying, or of being in a strange
environment, cause the eyes to move in a myopic (nearsighted) direction. It was his philosopny that in an atmosphere of increasing anxiety myopia becomes epidemic, a contagious disease. The causes, then, as Bates saw them, are holistic, and they relate to effort and strain, mental habits which influence tne perceptual and
anatomical functioning of the eyes.
DEVELOPMENT OF THE MODEL
A description of recent neurophyslological evidence follows which provides a model for underatending the mechanisms which
underlie Bates’ observations. The literature of neurology has continued to add new information about the process or vision. The technology of scientific inquiry has allowed us to get closer to some basic aspects of brain and muscle function than was possible fifty years ago. Perhaps Bates’ evidence would receive greater substantiation in todayfs scientific community.
If mechanisms are found which explain how mental strain can cause the extraocular muscles to change the refractive state of the eye, and if the model can relate to the literature reviewed in the first section of this chapter, the questions of remediation and prevention would be more accessible to future scientific study. Such a model would also help to extabllah Bates’
credibility with the professional community and might also change the present paradigm of refractive treatment.
The scientist whose work appears to have the most relevance to this topic is Karl Pribram. His contributions to neurological thinking attempt to amalgamate the fields of neurology, physiology, and the behavioral sciences. The model developed in this section relies heavily upon his book Languagesof the Brain (Pribram, 1971).
Pribram’s approach to neurophysiology Is unique in the field. In fact, he calls his area of research “neuropsychology” (Pribram, 1971) which is the multidlsclpllnary approach to research.
…my experiments, to a large extent, deal with determining by behavioral analysis the function of various systems of neural structures that make up the brain. This “systems” neuropsychology furnishes a halfway house between neurophysiology: the electrical and chemical study of the functions of nerve cells (and their parts), and experimental psychology: the behavioral analysis of functions of the organism-as-a-whole.
The systems neuropsychologist perforce, therefore, listens to – and relates hia investigations to – disciplines that have encountered their own concepts and styles to deal with these problems.
Pribram (1975) calls for a more comprehensive view of
psychological processes to establish a system of what he calls “scientific psychology” which concerns problems of
consciousness, perception, imagining and attention.
…paradox was encountered in experimental results that puzzled because they departed from those
predicted and thus made suspect the predictive value of currently held views about how the brain was supposed to work. So new theories and theses were developed. (Pribram, 1971)
At this stage, the theory must of necessity be primarily inductive, relying on a systematization of available data and drawing upon metaphor and analogy from more advanced knowledge concerning other physical, biological and social
organizations for initial model construction, (Pribram, 1975)
Pribram is considered a pioneer in this area, and has been suggested as a possible Einstein of an emerging paradigm of consciousness (Ferguson, 1977). His grander ideas concerning the merging of “spirit and science” are beyond the scope of the dissertation, his
research and theory encompass the whole spectrum of human consciousness: learning and learning disorders, imagination, meaning,
perception, intention, paradoxes of brain function. (p. 2)
The new theory has awesome implications in terms or the individual’s potential to affect his life — his reality — and impressive power to unify disparate discoveries in consciousness research. (p. 3)
Pribram’s theory has gained increasing support and has not been seriously challenged. (p. 3)
This dissertation has drawn from only a small portion of Pribram’s work. An attempt has been made to include
information which would be accepted by more moderate investigators in tne fields of brain and vision researon. The adaptation of systems neuropsychology or scientific psychology to the
practlcal problsma in optometry and ophthalmology has not been attempted before now with the possible exception or Bates’ (1921) and Huxley’s (1942) writing.
MENTAL CONTROL OF VISION
In order to more easily understand the Information presented in this chapter a descriptive model of sensory-motor organisation has been created. There are different ways an organism can organise itself in its interaction with the environment. Pernaps the following example will illustrate what is meant by sensory-motor organisations.
If we examine the normal behavior or a police force in a city, we see that the officers in squad cars (which are
analogous to the sensory receptor systems of an organism) travel around the city looking for meaningful situations. They look for people who are breaking laws – who are running red lights, fighting in bars, being disorderly, behaving obscenely, selling or using illegal drugs, speeding, etc. When such a situation is perceived, the parties are cited, arrested, reprimanded, or handled in some routine automatic manner. Information is
recorded and filed, usually reported to the police headquarters, and, in turn, recorded in some government computer. In addition, the computer might be searched for past records which could be meaningful to the current situation. The police chief and administrative staff are usually unconcerned or unaware of any specific details. They only care that the level of activity is proceeding normally. No analysis of the activity is carried out, and the police chief can be speaking to the Lions Club or the FTA, golfing, or having lunch with the major, or filling the abstract, higher order, or future needs of the police department.
But if one of the patrol officers ia wounded at the scene of a bank robbery, and has reported to headquarters that tne bandits have escaped in a red Ford automoDile, then the behavlor and organization of the police department cnanges rapidly. Calls go out to all available patrol cars; the chief is summoned from his dinner; the central radio control operators are alerted to attention and stand ready to relay information to the chief; news media people converge to the scene, etc. The patrol oars now turn on their sirens and rush to tne bank, ignoring any petty misdemeanors wnicn occur along the way. The information of interest becomes all important as all the police look for the same thing: the telltale red Ford. At each sighting of a red Ford the information is transmitted to headquarters and then to the chief who analyses the input, plots it on a map, and alerts and directs the other patrol members according to the emerging pattern. This feedback loop is continued until the pattern becomes predictive and a feedforward plan is plotted out on the central map: the red Ford is going west on Highway 1, will arrive at the bridge in nine minutes, patrol cars and officers are directed to seal off all exits to block off the bridge, and a helicopter covers the scene until the offenders are apprehended. The all-clear is given and patrol cars are again on the lookout for shoplifters, flashers, and student protesters. The novel episode has been properly integrated into the records of the police files, and the police chief can return to hia dinner or playing cards with the newspaper reporters.
This is an example of the simple reciprocal operational
organization of a peripheral-central sensory-motor response system. If the red Ford silently disappeared or if the bandits took over a school bus full of children as hostages, attached and
commandeered a police car, or went to the local airport with a large bomb, the process would become more complicated and pressures within the department would be greater. The officers on patrol would become more nervous and trigger-happy and the chief might lose his job. As long as the situation persists, the department cannot return to normal functioning, yet neither can it maintain its vigilant state indefinitely. Thus the distinction between the two modes of police organisation becomes less clear, with the result that the system comes under increased pressure to develop new levels of control to cope with the confusion and uncertainty of the situation.
The brain is constantly reorganising itself according to its needs and according to forces in the external world. In the above modelt the police officers in their squad cars
correspond to the sensory receptors. The police radios represent the neurological transmitting elements which communicate
information from the receptors to the appropriate brain stations and from the brain to the receptors. There is two-way communication. The police headquarters and police chief correspond to the brain.
In dealing with the situation brought about by the bank robbery and wounding of the police officer, the process of
reorganising the various parts of the police force represents a
model of how individuals can reorganize themselves. An extreme event can alert the brain to bring about changes in sensory
organization. The organism attends only to pertinent aspects of the environment; it ignores the rest. Instead of merely
receivIng and recording data from the outside world, the brain begins to reason and to plan, and to control the action of the total organism in more goal-dependent ways. If the situation resolves itself quickly, the system recovers its original organisation. If the situation persists, the organism continues to alter its function, constantly changing, vigilantly directing and
controlling its action according to new data. A sufficiently stressful situation, or one prolonged for too long, can bring about chronic changes in the organism (Pribram, 1971; Bates, 1920).
Neurophysiological mechanisms and models have been
discovered and conceptualized which can account for such behavior in humans. There is evidence that brain centers have direct
control over visual input even at the retinal level.
CENTRAL CONTROL OF VISUAL PROCESS
Evidence describing central control over sensory input has been produced by several experiments (Pribram, 1971). In one experiment a tiny electrode is inserted into a lateral
geniculate nerve cell. The lateral geniculate is located in the
subcortical brain and is the next step in the visual processing pathway after the retina. The electrode measures tiny
electrical nerve discharges in the nerve cell as it responds to visual
stimulation through the eye. By presenting visual targets at various places in the visual field, the response characteristics of the nerve cell can be measured and mapped. In some locations the visual targets will cause the cell to alter its electrical discharge rate, in other locations there is no change in
discharge rate.
In the geniculate and retinal cell experiments the
receptive fields are generally round in shape and usually have a
central area which, when stimulated, causes the cell to fire more rapidly. Surrounding this central area is a peripheral area which inhibits the firing of the cell. Such receptive fields are said to have an on-center and an off-surround. Pribram {1971, 1974) found that the size of the central area of the receptive field did not always stay the same. The size of the central area changes if specific brain areas are electrically stimulated while a receptive field is being mapped for a given nerve cell in the lateral geniculate.
Two areas of cerebral cortex have been shown to influence the neural organisation of visual input (Pribram, 1971). One is in the inferior posterior temporal lobe cortex (also known as the visual association area). The other is in the frontal lobe cortex (also known as the frontal, frontal cortex, or frontal llmbic). See Figure 6. If the visual association
cortex is electrically stimulated, the receptive field central area measures markedly smaller than when there was no
stimulation of cortical brain areas. If the frontal cortex is
stimulated instead, and the receptive field is again replotted, the
reverse effect is recorded; the receptive field center now plots larger than before. The size of the visual-spacial organ!zation is changed by stimulating selected areas of the brains cortex.
Evidence for central influence on the retinal organization la obtained when the intensity of electrical discharges in the optic nerve is compared during stimulation of these same brain centers. The discharges measured in the optic nerve are visual responses to two light flashes which stimulate the retina. One light flash is followed quickly by the second. If the
electrical nerve discharge to the second flash is less than it was for the first flash, we know that the second response was affected by the first and that the retina had not recovered from the effects of the first flash. When the association cortex is stimulated and the experiment is run, it takes a longer time for the retina to recover from the first flash (ie., the flashes have to be separated by a longer time interval for equal discharge), and a shorter time when tne frontal cortex is
stimulated (Pribram, 1971). This is direct evidence of tne two-part
control that tne brain has over tne visual system at the retinal level. In addition, non-visual input (such as tapping the paw or auditory clicks) cnanged the electrical activity of the retina in cats (Pribram, 1971).
BRAIN ORGANIZATION FOR TWO TYPES OF FOCUSING ATTENTION
If a rabbit notices a potential predator, it will become alert, set up an internal plan of action (flight) and implement the plan. If its response pattern is intact, it will run quickly
to cover and will ignore any appetizing berries it comes across along its path. It thus selects which aspects of the environment to attend—the internal plan, or the external stimulation. Once it reaches safety and the danger has passed, it will again search the environment for potential food, sex, fight, or flight stimulation. Thus, it can selectively open itself up to the
external environment or close itself down.
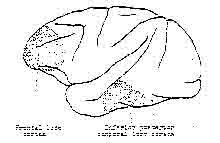
Figure 6 – Schematic cross section of the brain.
Shows the approximate locations of tiie frontal lobe cortex and the inferior posterior temporal lobe cortex. Based on Pribram, 1971.
POSTERIOR ORGANIZATION
The brain is essentially a decision-making mechanism. The organism must be aware of complexities of input which lead to actions. It must attend to information relevant to decisions. It must recognize the familiar and distinguish the novel. The association areas of the posterior cortex, in conjunction with lower brain centers, are involved with search processes. It is involved in organising and preparing for motor actions such as voluntary saocadic eye movement (see definition of terms}. It attends to overall, broad perspective input and is concerned with making decisions about what to do about what is perceived (Pribram, 1971). Its action is analogous to the routine
situation in the police department model. Police are moving about the total environment searching, acting, and handling isolated events without much direction from headquarters. The situations are routine; they are recorded in the log book and then forgotten.
This is the function of the association complex of
organization: making matches between outside events and long-term knowledge, seeing much, but not stopping to analyze deeply what
is there. There is no episodic sense or temporal continuity, Each event is a relatively isolated, acted upon, and then
dismissed experience. In reading, for example, some children are very adept at naming the letters or calling the words but at the end of a paragraph they cannot tell you what they just read. There is no sense of continuity, no context, just a series of unrelated eventa matched to long-term memory. A person can flow along, moving from one experience to another, attaching no
meaning to the experience. This seems to be the style of the
association are (Pribram, 1971).
FRONTAL ORGANIZATION
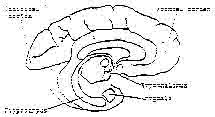
Figure 7 – Schematic cross section of the brain.
Shows the locations of the hypothalamus, amygdala, and the hippocampus.
The frontal cortex and its lower brain connection serve a different function. The frontal system centers on specific
details in the environment. It organises sensory input in order to categorize, record, and identify surprising or unique input. It shuts down awareness of the larger aspects of the environment in order to concentrate on and identify smaller details. It provides a context and a meaning for the input.
When the bandit robbed the bank and shot the officer, the police headquarters was alerted and the police chief was aroused. The police chief is analogous to the frontal cortex. The chief alerted the patrol officers and altered their action, gathered and organised the incoming Information and finally created a context for meaningful action. The frontal system seems to be involved in arousal and discrimination tasks; it works to make the unfamiliar familiar. It closes the organism down with
to awareness or the greater environment, and it curtails motor action (Pribram, 1971) . Thus, the mental system of
apprehending the world and acting on it is seen to be a product of two major brain organizing systems — the frontal and the
posterior — and a third system in the limbic area of the brain, the Hippocampus, which controls and coordinates the other two, Figure 7. These changes in configuration of brain organization include the peripheral sense organs themselves.
CENTRAL CONTROL OF VISUAL ACUITY
In the case or the organization of the eye, the size of the central area of rsoeptive fields recorded from retinal ganglion cells changes depending upon cortical stimulation. Frontal stimulation causes an expansion, and posterior temporal stimulation a shrinking of the receptive field center. These changes in spacial organization are due to lateral inhibition or contrast enhancing mechanisms which are abundant in the nervous system (Pribram, 1971}. The area of a receptive field is large compared with the area of a photoreceptor (rod or cone); individual rods and cones connect with many ganglion cells and each ganglion cell receives connections from many rods and/or cones, Figure 8. Any given area of retina does not belong to any one particular ganglion cell but instead, many ganglion receptive fields overlap the area. The receptive field center is surrounded by an inhibitory border area (i.e. instead of causing the ganglion cell to fire more often, stimulation of this portion of the receptive field results in a decreased
firing well below the spontaneous rate for that ganglion cell) (Pribram, 1971). Stimulation of one area of the retina leads to inhibitory interactions among adjacent neurons (ganglion cells). This facilitates the sharpening of spatial patterns.
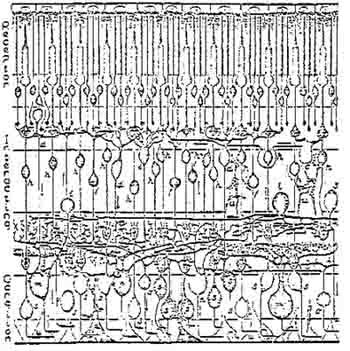
Figure 8. Scheme of the primate retina.
Rods and cones connect with many ganglion cells and each ganglion cell receives
connections from many receptors. From Polyak, 1941.
Activation of the frontal brain organization acts to increase the contrast between adjacent areas of the field. This increases the resolution of details and, thereby, increases the acuity of the visual system. Posterior association cortex activation works in a reciprocal manner; resolution of detail is decreased and, therefore, visual acuity is reduced.
Changes in visual acuity are thus dependent upon brain and retinal states as well as on the focus characteristics of the optical system of the eye. The frontal and posterior reciprocity and its effect upon contrast enhancing mechanisms provides a much needed explanation for the variability in the relationship between visual acuity and refractive error. It also explains how acuity can improve without a corresponding change in refractive error as reported by Kelley (1958), Zeiger (1976) and others.
In summary, Bates (1920) suggested that the underlying cause for refractive error was mental strain or loss of mental control and clarity which, in turn, is caused by emotional, intellectual, and social factors. The information from the neurophysiological studies presented above provides a basis for understanding what might be happening in the brain and sensory systems and tends to support Bates’ seventy-year-old concepts. The above information also provides a neuropsychological
explanation for much of the psychological literature on nearsightedness.
EMOTIONS, MENTAL STRAIN, AND VISION
Kelley (1958) found that the threat of electrical shock created a myopic response in eight out of ten normally emmetropic boys. Bates (1920) reported an instance of a girl whose myopia increased while she was telling a lie. Others (see Chapter Two) have reported some chronic personality and anxiety patterns correlated with myopia. This section will describe this relationship of vision to emotion in neurological terms.
THE AROUSAL SYSTEM
The frontal system of neurosensory organization works in conjunction with other areas of the brain. These include the amygdala and the reticular system of the hypothalmus and brain stem (Pribram, 1971). This frontal system is intimately related to the arousal-orienting response reactions of the organism. When an animal is alerted by a novel situation the frontal system is engaged and the visceral-autonomic centers in the core brain are stimulated. Changes occur in breathing, heart rate, blood flow, pupil size, skin resistance, etc., and the motor system orients (turns towards the action and then freezes). There are also changes in electro-encephalographic measurements – brain wave rhythms increase in frequency and become desynchronized. If the stimulus is especially strong, surprising, or appears especially dangerous, the reactions of fight or flight
take place.
In the example of the police department, the novel situation stimulated the wounded officer to alert headquarters and arouse the police chief, which action, in turn, changed the sensitivity and reaction of the entire police force. In the brain, novel stimulation changes brain function in an analogous way through the arousal system. The amydgala and hypothalmic structures are the control and activation centers of the arousal response. If the frontal cortex and/or the amygdala is eliminated from the brain (through accidental brain trauma or through experimental tampering) the visceral-autonomic aspects of the arousal response disappear, but the behavioral-motor and electro-encephalic reactions remain intact, often continuing to react for longer periods of time than under normal, intact brain conditions (Pribram, 1971). The amygdala and hypothalamus are commonly known to be centers for emotional reactions (Bard, 1961) (See Figure 7). Thus the brain organizes itself according to the stimulus-response demands of the situation and part of this reorganization involves the emotional-motivational centers of the brain.
MOTIVATION AND EMOTION
According to Pribram’s (1971) model, motivation and emotion are reciprocally connected and are expressions of the relationship between perception and action. When information is familiar to the organism it is immediately meaningful and is recognized and acted upon without effort expended to classify
or understand it. The response pattern in this case is mainly motor (action) with little need for the autonomic changes which an especially novel environment stimulates through the arousal reaction.
If the information is not familiar and is confusing, threatening, or overwhelming to the organism, a different brain process is engaged which includes arousal, frontal organization and vigilance (i.e. the information will be processed until it is understood).
Suppose the rabbit, in our previous example, were involved in an emergency wherein the environmental stimulation overwhelmed the rabbit’s ability to act in a meaningful way. For example, if a dog came running towards the rabbit from the left, the rabbit would alert, go into its flight reaction and run for cover to the right (motivation leading to action). If another dog suddenly appeared coming from this new direction, the rabbit would freeze. It would be at a loss to make a decision about where to run (emotion and inaction). It could make a break for possible cover in a new direction or it could remain frozen in position. But the situation would no longer be automatic and the rabbit would have to organize itself to cope with its uncertainty.
According to Pribram (1971), the organism has two ways to internally adjust to its uncertainty. It can increase the rate at which it processes information (frontal organization) or it can decrease that rate (posterior organizing action). It can cope by opening to external input through enhancing its monitoring
of the external environment, or it can restrict input by minimizing the external and focusing on internal configurations. The first way implies motion or motivation, the second emotion. When the variety of perceptions exceeds to some considerable extent the repertory of action available to the organism, it feels “interested” and is motivated to (i.e. attempts to) extend this repertory. Whenever this attempt fails, is non-reinforced, frustrated or interrupted, the organism feels emotional, (i.e. the coping mechanisms of self-regulation and self-control come into play). Furthermore, emotion is likely to occur when the probability of reinforcement from action is deemed low. Pribram (1971) states:
The suggestion is that ordinarily interests, feelings of motivation and emotion, occur when the organism attempts to extend his control to the limits of what he perceives. To the extent that this attempt appears (on the basis of trials or experience) feasible at any moment, the organism is motivated; to the extent that the attempt appears infeasible at any moment, the organism becomes, of necessity, emotional. (p. 212)
Since the frontal system is so intimately involved with the arousal system, and the arousal system is an autonomic-visceral brain system dependent upon the homeostatic balances in the body., it is in this system that we have such a close tie with emotions and nutritional and metabolic balance.
VIGILANT ATTENTION
An area of the forebrain called the hippocampus seems to be responsible for controlling or prolonging attention (Pribram
and McGuinness, 1975). The ability to voluntarily prolong
states of attention is interfered with when the hippocampus is lesioned. In experimental animals, with hippocampus lesions, voluntary control over brain organization decreases. They are left at the mercy of involuntary reactions, and either sensory input or motor output dominates behavior. The voluntary control operations are absent (Pribram and McGuinness, 1975).
In higher order tasks, such as categorizing and reasoning, a prolongation of the states of arousal and readiness for action is maintained voluntarily to allow for the internal rehearsal necessary to organize the perception or response pattern. Such tasks are called vigilance tasks and they require the voluntary disruption of the stimulus-response pathways in the brain organization. These tasks might be termed the “What is it?” (categorizing) and the “What is to be done?” (reasoning) modes of behavior. In the case of the rabbit, for example, when the predator was noticed, the defense reaction was fully automatic, the sensory organization closed down to external cues and focused upon internal plans of action.
CATEGORIZING TASKS AND MOVEMENT
In categorization there are autonomic as well as muscular changes in behavior. In studies of heart rate and muscular movement (Pribram and McGuinness, 1975) ability of children to match photographic slides from memory (vigilant attention to categorize) was accompanied by heart decleration and decreased motor response. The longer and more decelerated the heart rate, and the more the reduction of movement, the more likely the children were to make
a correct match. A similar relationship was found with I.Q. and heart and muscle changes. The ability for the brain to organize itself for this skill (vigilant attention for categorization accompanied by autonomic and motor changes) seems to be developmental and occurs earlier in girls than in boys. The ability increases linearly with age for both sexes. This means that the ability to maintain attention to outside stimuli, while at the same time retaining awareness of recently perceived events which are now stored in memory (short-term memory) is related to a brain organization which produces a decrease in heart rate, an inhibition of motor activity (including the muscles of breathing) and other physiological changes.
When the organism prolongs attention to the complexity of input in a categorizing discrimination task, he reduces extraneous noise by eliminating random movement. Heart rate then adjusts to the reduced demands of the system reflecting characteristics of the somatomotor system (Pribram and McGuinness, 1975)
REASONING TASKS AND ISOMETRIC CONTRACTION
Reasoning also requires vigilant attention but it is not the same as categorizing. In categorization processes, external information is attended for prolonged periods of time. Arousal precedes action. Reasoning, on the other hand, seems to be more involved with processing alternatives for action; motor readiness for action is internally rehearsed until a solution is found and then the action takes place (either externally as behavior, or internally as imagery or imagination)
which is now monitored by the sensory apparatus. In reasoning tasks, action precedes arousal. Categorization, the discriminating and registering of small details of the external world, seems to be related to frontal organization and arousal; reasoning (internally rehearsed alternatives to action) appears related to the posterior cortex control systems.
Reasoning seems to involve the activation of a special form of muscular and metabolic activity related to sympathetic nervous control. Oxygen uptake in the muscles and blood flow to the muscles decreases to much lower levels than in usual somatomotor activity. Muscle force increases but “the muscles ‘work’ under the special conditions of isometric contraction” (Pribram and McGuinness.. 1975). An example of this is the common practice of holding the eyebrows close together in an isometric contraction during reasoning tasks. In addition, optometrists are aware of changes in the eyeball during reasoning tasks such as math problems (e.g. Apell and Lowry, 1959).
COGNITIVE STATES AND REFRACTIVE ERROR
Gesell (1949) provides an in depth description of changes in refraction (including myopic, hyperopic, and astigmatic variations) and retinal characteristics (the color and brightness of light reflected off the retina) observed during categorizing and reasoning behavior. These changes were observed during retinascopic examination of children while performing various tasks. They (Gesell, et.al., 1949) note that:
The variations which are observed in a single session in a single child are not random, but are correlated with variations in his visual acts …. The reflex showed a definite against (myopic) motion when the child fixated the examiner’s race. Similarly, the reflex brightened and showed an against motion when an interesting toy was proffered. As the infant’s hand came in, there was a with (hyperopic) motion, followed by brightening as the hand grasped tone toy. During tactile-visual exploration of the toy, there was a succession of against, with, and against motions. The slowness of the motion suggested a wide range of oculo-cortical manipulations. (P. 176-177)
They further state:
The eye, like the hand (to say nothing of the mouth), is an organ of manipulation, and in higher adjustments each is separately, and both are conjointly, governed by the cortical brain. In the last analysis, it is the brain which appropriates the outer world, whether manually, orally, or ocularly. It does so by a projective process, but this is a two-way, reciprocating process – a directional process which emanates from within the self, goes out, and then returns within. Thus projection both externalizes and internalizes. These two distinguishable phases have their counterparts in the plus-minus (myopic-hyperopic) and minus-plus adjustments of the oculo-manual presensory apparatus. Skeletal, visceral, and cortical factors are simultaneously involved, but the manifestations are not fortuitous. They are lawfully correlated with the functional and maturity status of the action system. (P. 177)
It is this author’s contention that the retinoscopic changes observed and described above are due to the isometric muscular and the metabolic changes which occur during categorizing and reasoning tasks. This is certainly in concert with Bates’ ideas about the development of myopia (and other visual problems), which Bates (1920) related to peripheral manifestations of mental strain: “Every thought of effort in the mind, of whatever sort, transmits a motor impulse to the eye; and every such impulse causes a deviation from the normal in
the shape of the eyeball.” Bates held that refractive problems are due to habits of thought which develop as a result of the unhealthy conditions imposed upon individuals in educational and vocational activity. The cure, he felt, must reside in finding ways to change these faulty habits resulting in loss of mental control and poor vision. Bates (1920) claimed that even the circulation of the blood in the eyes and other parts of the brain was influenced by habits of thought. Confirmation of this idea has been indicated by studies of local differences in cerebral blood flow related to effortful problem solving behavior (Pribram and McGuinness, 1975). According to the model of brain organization described above:
…physiological (including muscular) responding during stimulus intake will depend entirely upon how a task is constructed and whether it is primarily a categorizing or reasoning problem (Pribram and McGuinness, 1975).
In situations where attention is maintained (categorizing) while an organism is involved with problem solving (reasoning), metabolic and muscular changes occur. If these conditions are maintained for long periods of time, they may become structured into the response patterns of the organism. The task variables which affect arousal and activation, and the effort required to retain information during categorizing and reasoning include the information content of the relevant stimuli and the degree of concentration required, i.e., the “cognitive effort.” Cognitive effort is defined as the
measure of attention paid to increase or maintain the efficiency of a communication channel (Pribram and McGuinness, 1975). This cognitive effort arises through voluntary restrictions placed upon one’s response patterns in an attempt to obtain a goal.
The reading task which has so often been related to myopia (Chapter II) certainly falls into the category of skills which require simultaneous categorization and reasoning processing. The reader must search and discriminate words and letters, register the information, digest its meaning and at the same time vigorously forstall the fading of recently acquired information (delay closure). Retinoscopic fluctuations in brightness and color of the light reflected from the retina, and changes in refraction in myopic and hyperopic directions and in astigmatism accompany the level of interest or disinterest, familiarity or unfamiliarity, and complexity of simplicity of the reading process (Getman, 1960). Myopic changes are commensurate with interested reading of unfamiliar or complex material.
Interacting with the everyday environment in many ways is similar to reading, and research exists which supports the notion that myopes are involved in a continual effort to see and to organize their perception of the world. Stimulation from the environment is categorized (what is it?) and actions are rehearsed (what is to be done?) for most people most of the time. But when an individual has high self-expectation and is motivated to act “correctly” (“…myopes are good children,” Kelley, 1971) and to achieve high success (Young, 1967) the
world-out-there must be read constantly and with great cognitive effort in order to conform to the self-directed (and externally imposed) goals. This is especially true when the goals are unrealistic.
EYE MOVEMENT AND MENTAL STRAIN
Bates observed the characteristic eye movement behavior of people with good, natural vision and compared it to the behavior of people whose vision is poor. He found that those with poor vision tend to hold their eyes in one position for prolonged periods.
If an eye with poor vision cannot see a particular point it stays there trying to make it out by staring at it (Bates, 1920).
An eye with good vision will shift its fixation, moving around looking from point to point until enough of the context is known for the image to become clear. It does not try to bring out the detail by staring at it, 11 … which is what the eye with poor vision does all the time (Bates, 1920).
The eye movement patterns characteristic of poor vision are similar to those of experimental subjects who are involved in situations of visual overload. Mackworth (1976) used sophisticated eye movement measuring devices to record the visual characteristics of subjects in visual tasks at various levels of difficulty. He described the adjustments made by the visual motor system to conditions of visual overload. The major adjustment was seen to be a narrowing of the size of the useful field of view. This was true not only during the active search of too much material in too short a time, but was also true during
tasks requiring vigilance, or waiting for a visual stimulus
event to occur.
He also found that the size of movements was reduced and more stereotypical. Despite the decrease in useful field
of view, the subject tried to include more stimuli in each fixation. The cognitive difficulty required to process
complex visual material was indicated by longer visual fixation times and attempts to attend to more items during each fixation. Finally, there was a complete breakdown as task complexity increased to conditions of visual overload.
We would expect that as the apparent complexity of environmental demands exceeded visual ability, visual-motor behavior would become habitually stressed as described by
Bates (1920), measured by Mackworth (1976) and predicted by Pribram’s model described herein.
Mackworth (1976) supports the idea that people have differing ways of viewing the world. He states that the useful
field of view (the amount of peripheral vision available to an
individual) varies in size between different types of tasks as
well as between different kinds of people. Some people can
attend to specific details and then quickly take a broader view
when contextual understanding is important while others seem caught up in an inflexible, unvarying perceptual style in any situation.
The brain initiates the physiological mechanisms of attention rather than waits for the environmental input to set these mechanisms in action. The interaction between brain and the environment is two-way, and the set or motivation of
the subject is as important as the physical input in determining the physiological responses. For instance, the pupil contracts reflexly when light falls on it, but it also changes according to the cognitive state of the brain. Similarly, the useful field of view sharply constricts when there is high density of detail to be processed by eye and brain. Both behavioral changes are attempts to prevent the processing mechanisms from being overloaded. (Mackworth, 1976)
MYOPIA, POSTURE, AND COGNITIVE EFFORT
Prolonged and chronic involvement in such tasks as reading, or situations which are demanding but difficult to understand (e.g., family or peer group situations in which there is a demand to act “correctly” in conformity to a reality structure which the individual does not understand) can lead to chronic isometric muscle patterns and metabolic changes. It is the thesis of this paper that myopia is one possible outcome of this behavior. The situation of a rabbit frozen between two charging, barking dogs if repeated for a period of weeks or months would likely lead to isometric muscle contraction and metabolic changes in the rabbit which become structured into the brain and body.
Humans exhibit chronic isometric muscle contraction patterns in various places in their bodies even to the extent that they grossly distort their physical postures. Lowen (1968) contends that severe chronic isometric constriction patterns are evident in the posture adopted by the majority of people in our culture. Darwin (1965) classified emotional expression (and suppression) according to isometric muscle contraction patterns. Reich (1949)
analyzed character and personality types an the basis of patterns of postural contraction. Alexander (1969), Feldenkrais (1972). Lowen (1958). and Reich (1949) have developed forms of psychomotor therapy to deal with this very type of muscle contraction. Lowen, who studied with Reich and later developed his own form of therapy called bioenergetics, has found myopia to be so common among his patients that the considers it abnormal if they are not myopia. With regard to his experience and observations Lowen (1964) states:
In my opinion myopia is a functional eye disorder that has become structured in the body as a distortion of the eyeball. It does not differ from other bodily distortions that are the result of chronic muscular tensions. In many cases these distortions are significantly reduced as the tensions are released. I have seen considerable changes produced in people’s eyes by bioenergenetic exercises and therapy. And I know one person who completely overcame his myopic condition through the Bates method. One of the difficulties in working with the myopic eyes is that the tense ocular muscles are not accessible to palpation and pressure. (p. 11)
Bates (1920) attributed myopia directly to isometric contraction in the extraocular muscles. These are the muscles which attach to the outside of the eyeball and are considered to be responsible for moving the eye in its socket (Alpern, 1962). The extraocular muscles comprise the physical mechanism which is responsible for manipulating eyeball length. If the oblique muscles of the eyeball (Figure 9) both contract simultaneously they can increase the cornea-retinal distance. The focus of an optical system such as the eye depends upon the refractive power of optical elements in the system, and it depends upon the distance of the optical elements from the
focusing screen (the retina in the eye). If the corneal-retinal distance increases too much, the rays of light entering the eye will focus between the cornea and the retina and not on the retina itself. This condition is defined as myopia. This is perhaps Bates’ most controversial concept.
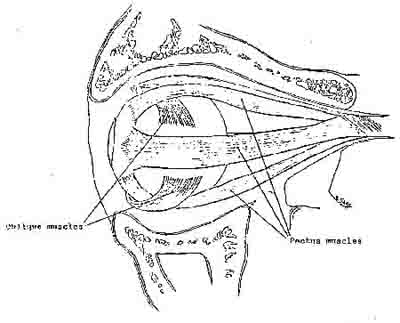
Figure 9. Side view of the eyeball and its extraocular muscles.
The general dicta of physiological optics only ascribes a movement function to the extraocular muscles and rejects the idea that they might also be responsible for changes in eyeball length (Huxley, 1942). Physiological optics does not conceive that the process of emmetropization involves active mechanisms which coordinate the ocular components of refraction, regulated by extraocular isometric contraction patterns involving oblique and rectus muscles.
What is the evidence which supports this idea? Bates (1920) reports experiments which he conducted on fish and mammals (including humans) which led him to believe that the extraocular muscles were responsible for refractive changes. In many varieties of sea creatures, it is known that the extraocular muscles are responsible for changing the focus of the eyes dynamically to see clearly objects at near and far distances. The focus of the fish’s eye is controlled by the relative tension of the oblique muscles in comparison to the rectus muscles. The same mechanism was found to exist in rabbits, cats, dogs, and humans (Bates, 1920).
Further evidence that pressures on the globe of the eye can produce changes in eyeball length, leading to refractive changes, exists in recent literature:
Succinylcholine-induced co-contracture of the four rectus and two oblique muscles in cats produced enophthalmos and up to 0.4 mm of globe-shortening as measured by strain gauges. Concomitant hyperopic refractive changes of up to 2 diopters were also noted in cats, and similar hyperopia refractive changes were noted in two human subjects (Bach-y-Rita, 1968).
Two types of muscle fibers and control systems exist in
the extraocular muscles. These muscle systems are “quite distinct motor systems”, one quick acting, the other more primitive and slower to act. The quick acting system is responsible for “rapid and wide-movements of the-eyeball, while the tonic system provides for slow, small movements and fixations” (Matyushkin, 1964)- Succinylcholine is related to sympathetic neural transmitter chemicals which Pribram and McGuinness (1975) have implicated in isometric and metabolic alternation of muscle action during cognitive effort in reasoning tasks. The experiment by Bach-y-Rita, et.al. quoted above, is similar to those reported by Bates (1920); both arrived at similar conclusions.
Additional evidence that pressure around the equator of the eyeball can lead to myopic changes comes from the inadvertent discovery
… that, when silicone bands are placed about the equator and tightened to reduce vitreous traction in retinal detachment work, the eyeball becomes longer axially and thereby increases myopia…. (Rubin, 1966)
Tentative support for extraocular muscle influence on refractive error was suggested by Fry (1966) who recommended that the tension on the eye by the tonus of the
extraocular muscles should be investigated as a possible cause of refractive error; and by Ogle (1966) who advised that studies be made as to the possible effects on the corneal curvatures of surgical procedures on the extraocular muscles.
Van Alphen (1966) proposed that the eye is self-adjusting, focuses itself to emmetropia, through cortical and subcortical control of the tonus or ocular muscles, and that ametropia is produced by factors which interfere with that mechanism. Ametropization as well as emmetropization may be active, ongoing processes in brain organization. He also stated his belief that the ocular muscular control centers in the brain stem and the reticular activating system are so intimately related that “psychological factors, stress and emotion, and extreme autonomic endowment” might very well affect the activity of these centers. He (Van Alphen, 1966) stated:
I have found some evidence for psychological differences between emmetropes and myopes, including indications that psychological factors may facilitate the development of myopia and not vice versa: I have emphasized the stress situation in the socalled school myopia; and I have also the clinical impression that many myopic children are distinctly nervous, If such factors interfere with the emmetropization mechanism at a subcortical level, one might expect imbalance in the ametropic eye. indeed, there are differences in pupil size in myopes and hypermetropes …. Admittedly, the extraocular muscles may well contribute to the origin of ametropia. (P. 31)
SUMMARY OF THE MODEL
A neurological, neuropsychological model has been presented which supports the notion that myopia results from habits of mental focusing (habitual ways of organizing mental process in
order to pay attention) which can lead to chronic isometric contraction of the extraocular muscles which lengthen the eyeball and lead to myopia. Voluntary attention involves mechanisms which prolong readiness for action and discrimination. The attention involved is voluntary in the sense that it is initiated by the organism rather than by some input event. Such tasks have been designated as categorization and reasoning. Categorization delays the process of closure (i.e. prolongs readiness for discrimination) when the stimulus situation is unfamiliar or confusing to the organism. Selective attention to external details must be maintained until enough appropriate information can be detected and categorized. Such processes involve the frontal and arousal systems of brain action. Reasoning involves the internal rehearsal of potential solutions (actions) just as categorization involves the internal rehearsal of potential perceptions. In reasoning tasks the response is delayed until the solution is internally experienced (e.g., in mental arithmetic or in playing chess). Reasoning demands the uncoupling of attention from the immediate input variables and involves the posterior temporal (association) cortex.
Special forms of muscular effort involved in problem solving while categorizing or reasoning have been discussed. These have been implicated as the source of the chronic isometric muscle contractions linked to psychologically related postural distortions and to the extraocular muscle contraction which leads to myopia.
Electrophysiological evidence has been cited indicating that the posterior and frontal cortex contribute opposing controls on sensory channels. This evidence is based on changes produced in the recovery cycles of the visual system (the speed with which the system recovers to its full capacity after a sudden, intense stimulus) and the alterations produced in the shape of visual receptive fields. This has been implicated as a probable explanation for fluctuations in visual acuity which are known to exist in the absence of changes in refraction. Emotions occur as a result of involuntary self-control mechanisms triggered by environmental stimulation which is beyond the ability of the organism to deal with effectively. Mechanisms similar to those of categorization and reasoning are involved in emotional responses due to the decoupling of arousal and action. Emotional reaction implies inaction and can result from direct environmental stimulation or from projected inability to cope with potential environmental situations on the basis of previous failure or frustration in similar situations. Emotions are related to arousal and therefore to frontal system organization. The organism is motivated to seek pertinent information, in the stimulus situation, but is inhibited from doing so by the over-reaction of the arousal mechanisms. This causes the organism to close down prematurely from external stimulation by triggering emotional reactions (fight or flight).
…The organism’s nervous system is temporarily swamped by the arousing input and reacts defensively to shut out all further input and thus leads to automatisms (Pribram and McGuinness, 1975).
Mental strain prolongs muscular strain. Chronic involvement in emotional and/or categorizing-reasoning situations leads to muscular and metabolic changes in the organism. These become patterned into the mental response systems;
It is thus plausible that the neuromuscular processes demanding effort are to a large extent initially peripheral in their origin and manifestation. But as problem-solving progresses and problem-solving skill develops, the effort becomes more and more a concomitant of the brain processes involved-apart from, or only reflected in, the peripheral manifestations-and thus becomes truly cognitive (Pribram and McGuinness, 1975)
That is to say, they become habits of mental strain which manifest throughout the body, including the retina and the extraocular muscles. If an individual is continually involved in situations which are unclear or not easily resolved (searching while problem-solving), changes in sensory-motor, metabolic, and brain organization occur which can lead to myopia.
If an individual has been successful in coping in certain situations through internal control mechanisms (i.e., cognitive effort), the habit can become established even in less demanding situations. Darwin (1872) observed that:
Actions which have often been of direct or indirect service, under certain states of the mind, in order to gratify or relieve certain sensations, desires, etc., are still performed under analogous circumstances through mere habit although of no service. Even when these and other emotions or sensations are aroused in a very feeble manner, there will still be a tendency to similar actions, owing to the force of long-associated habit; and those actions which are least under voluntary control will generally be longest retained. (p. 81-82)
Actions which are least voluntary (such as co-contracture of the oblique extraocular muscles) may be altered when they are brought under voluntary control.
CONCLUSION OF THE CHAPTER
Bates (1920) concluded:
Primarily the strain to see is a strain of the mind and, as in all cases in which there is a strain of the mind, there is a loss of mental control. These facts appear sufficient to explain why visual acuity declines as civilization advances. Under the conditions of civilized life men’s minds are under continual strain. They have more things to worry them than uncivilized man had, and they are not obliged to keep cool and collected in order that they may see and do other things upon which existence depends. If he allowed himself to get nervous, primitive man was promptly eliminated; but civilized man survives and transmits his mental characteristics to posterity. (p. 98)
The etiological model which has been developed here serves
to integrate the work of Bates with recent psychophysiological findings and provides for the expansion of our notions or visual impairment and its possible cure.
What emerges here is a portrait of myopia as not merely a problem of refractive error but as a posture which the myope has adopted to deal with life.
It is the author’s clinical observation, supported by the current study, that myopia is caused by some crisis in reality wherein an individual is compulsively searching for clues, continually attempting to discriminate details of the environment in order to figure out how to act, to make the best choice, to achieve the most success, to get the most meaning out of the story, etc.
It is clearly indicated that the work of Bates, so long ignored by the scientific and clinical communities, needs to be reexamined and tested in an open-minded, scientific manner.
A comparison of this model with the evidence about myopia presented in the literature review and the conclusions and further implications derived from this chapter will be presented in the final Chapter of the dissertation. The following two chapters describe a pilot study of myopic patients.
1. This book is not generally available. Better Sight Without Glasses, published in 1940, (nine years after his death) is not the same as his 1920 book. It was reworked by his widow and lacks the magnitude of The Cure of Imperfect Sight Without Glasses.